时间:2022-05-25 15:00
内皮细胞(排列在血管管腔内表面的特化细胞)在神经系统功能中起着至关重要的作用。越来越多的证据表明,内皮细胞损伤是导致各种神经系统疾病的主要原因。
既往AAV血清型改造主要集中于:转导神经元或星形胶质细胞。相比之下,专门针对中枢神经系统内其他细胞群(包括内皮细胞)的AAV血清型相对较少,尽管人们开始认识到大量的非神经元细胞类型对神经系统功能至关重要。
迄今为止,只有两种有效转导脑血管内皮细胞的病毒载体被报道:AAV-PHP.V1[2]和AAV-BR1[3]。其中,AAV-PHP.V1在CNS中除了可以转导内皮细胞外,还高效标记星形胶质细胞。相比之下,AAV-BR1特异性较高,但其靶向脊髓和视网膜血管系统的能力是模糊的。
美国麻省理工学院-哈佛大学布罗德研究所的Benjamin E. Deverman研究组和哈佛医学院的Chenghua Gu研究组通过对AAV9血清型进行一系列改造,开发了一种新血清型AAV-BI30。并于近日在Nat Cardiovasc Res杂志发表[1]。
AAV-BI30这种新血清型能够特异且高效地转导整个中枢神经系统(CNS)的内皮细胞。在小鼠、大鼠模型和体外人类微血管内皮细胞中均能稳定表达。

图1 | DOI:10.1038/s44161-022-00046-4
AAV-BI30有效转导内皮细胞
首先,作者验证了AAV-BI30对内皮细胞的特异性。在C57BL/6小鼠静脉注射AAV-BI30,10d后皮层和海马中可见明显的内皮细胞标记,而较少转导外周器官微血管系统中的内皮细胞。在大鼠中也有同样的结果。
然而,在给药约1周后,观察到意外的剂量依赖性毒性(1×1011vg/只小鼠),表现为体重减轻、嗜睡行为等(如表中蓝线)。加入肝细胞特异性microRNA靶序列miR122后,这种毒性及肝脏富集的情况消失。
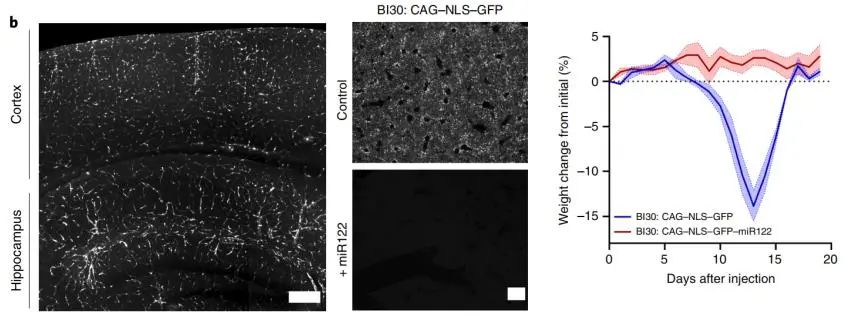
图2 | AAV-BI30标记小鼠皮层和海马内皮细胞(左),及miR122对肝脏富集/小鼠体重的影响
较AAV-BR1,AAV-BI30更优
那么这种血清型与AAV-BR1相比,是否具有优势呢?
作者发现,在相同的注射条件下,AAV-BI30标记的小鼠脑内皮细胞数量明显多于AAV-BR1。同时,AAV-BI30的表达在整个大脑中没有明显的区域间差异,即皮质、海马、丘脑和小脑均表现出>80%的内皮细胞转导。此外,与AAV-BR1不同的是,AAV-BI30具有高度内皮细胞特异性,标记神经元或星胶细胞的情况罕见。
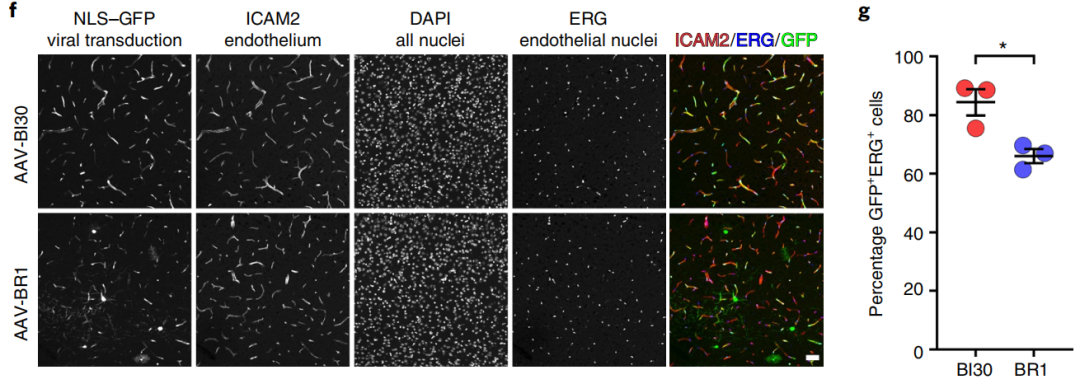
图3 | AAV-BI30与AAV-BR1标记情况的差异
除毛细血管外,AAV-BI30对动脉和静脉内皮细胞也有高特异性
毛细血管,这种大脑中最小的血管,构成了脑血管网络的绝大部分。但除此之外,在软脑膜中还分布着大动脉和静脉,这是在脑切片中难以观察到的。
为了观察AAV-BI30对这部分血管的标记情况,作者通过内皮特异性转录因子ERG、α-平滑肌肌动蛋白与GFP的共标识别。结果显示,AAV-BI30能够识别大部分动脉和静脉内皮细胞,这是在AAV-BR1未观察到的。值得注意的是,AAV-BI30对大脑最大动脉同样具有强大的特异性转导(结果未显示)。
这些结果都说明,即使在相对较低的剂量(1×1011vg)下,AAV-BI30仍能完成对全脑动脉、毛细血管和静脉内皮细胞高效且高特异性的标记。
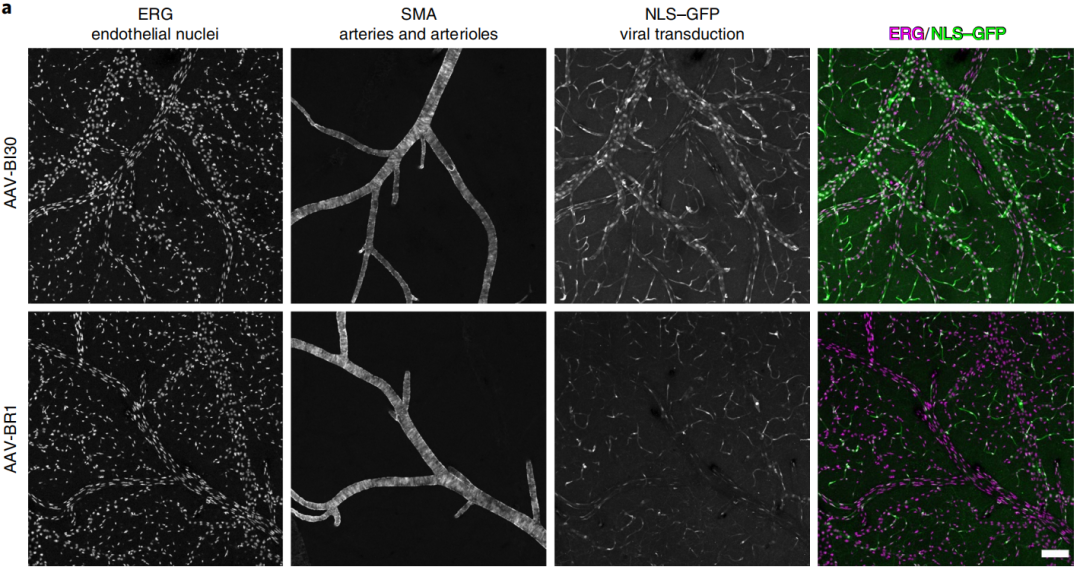
图4 | AAV-BI30和AAV-BR1在软脑膜血管中的转导差异
AAV-BI30还能转导视网膜和脊髓内皮细胞
虽然迄今为止,大多数内皮靶向的AAV研究都集中在脑血管系统上。但视网膜和脊髓血管系统也对血管生成、血脑屏障动力学、神经血管病理学和许多其他关键过程的研究至关重要。因此,作者还评估了AAV-BI30转导这些内皮细胞的能力。
图5所示,在视网膜的所有血管系统中,AAV-BI30显著优于AAV-BR1。在脊髓中也有类似的结果(结果未显示)。这说明,AAV-BI30的高效、高内皮细胞特异性嗜性并不局限于大脑;相反,它扩展到了整个中枢神经系统。

图5 | AAV-BI30视网膜内皮细胞标记效果显著优于AAV-BR1
AAV-BI30可用于脑内皮细胞特异性基因调控
为了评估AAV-BI30在体内对中枢神经系统内皮细胞的基因调控能力,作者将AAV-BI30:CAG-Cre-miR122-WPRE注入Rosa26:CAG-LSL-tdTomata(Ai9)小鼠,表达12d后,在整个脑微血管系统以及位于软脑膜表面的动脉和静脉中均观察到tdTomato的强表达。

图6 | 内皮细胞特异性基因调控
作者还用小窝蛋白(Caveolin-1)敲除的实验,验证了AAV-BI30介导的Cre递送的高度特异性。结果显示,小窝蛋白的缺失只出现在内皮细胞中,小动脉平滑肌细胞层中小窝蛋白表达则无明显改变。这些都说明,AAV-BI30介导的Cre递送具有高度特异性。
鉴于目前还没有中枢系统内皮细胞特异性的基因调控方案存在,AAV-BI30介导的Cre递送系统可能成为常用的CDH5:CreERT2或TIE2:Cre等系统的良好替代。
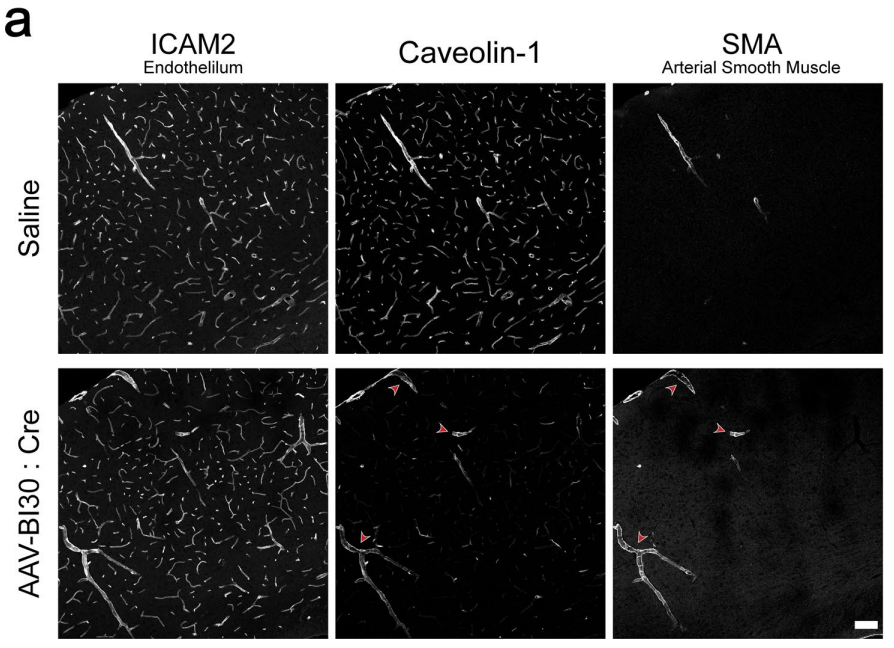
图7 | AAV-BI30介导的Cre传递实现了高效的内皮特异性基因缺失
总 结
AAV-BI30是一种极具潜力的新型血清型,它在CNS中的嗜性几乎只限于内皮细胞,且无需借助特异性启动子,因此具有广泛的可操纵空间,非常适合各种类型的应用。例如CNS内皮细胞示踪、细胞形态观察、血管网络重塑、功能调控、基因编辑、实时成像或双组分调节表达系统等。
这种血清型也非常适合加速我们对正常生理中神经血管相互作用的理解,解决神经血管疾病相关机理问题,并帮助开发治疗内皮功能障碍的治疗方法。



